Le ammoniti
-
Generalità
Le ammoniti (sottoclasse Ammonoidea) sono un gruppo di Molluschi cefalopodi estinti, comparsi nel Devoniano Inferiore (circa 400 milioni di anni fa) ed estintisi intorno al limite Cretaceo Superiore-Paleocene (65,5 ± 0,3 Ma) senza lasciare discendenti noti.
.jpg)
Si tratta di animali di ambiente marino, caratterizzati da una conchiglia esterna composta prevalentemente di carbonato di calcio, sotto forma di aragonite, e in parte di una sostanza organica di natura proteica (conchiolina). La conchiglia era suddivisa internamente da setti in diverse camere, di cui il mollusco occupava solo l'ultima (camera d'abitazione). Le altre, che componevano il fragmocono (parte concamerata della conchiglia), erano utilizzate come "camere d'aria" (analogamente all'attuale Nautilus), riempite di gas e liquido camerale per controllare il galleggiamento dell'organismo. La pressione dei fluidi camerali era controllata da una sottile struttura organica tubolare riccamente vascolarizzata, in parte mineralizzata (il sifone), che attraversava tutti i setti e permetteva lo scambio di fluidi dal sangue e dai tessuti molli dell'animale alle camere tramite un processo di osmosi. L'ammonite poteva così variare la propria profondità (entro i limiti di resistenza meccanica della conchiglia) in maniera simile ai nautiloidi tuttora viventi. Verosimilmente le ammoniti, come tutti i cefalopodi conosciuti, erano organismi carnivori, e secondo gli studi disponibili svilupparono probabilmente un grande numero di adattamenti diversi, dalla predazione attiva di animali marini, alla microfagia (predazione di microorganismi), alla necrofagia (consumo di carne di organismi morti), e persino al cannibalismo (predazione di altre ammoniti, anche conspecifiche).
Si trattava certamente di organismi con dimorfismo sessuale spesso marcato. L'ipotesi maggiormente accreditata è che i due sessi corrispondessero a morfotipi diversi di conchiglia: le femmine corrisponderebbero al morfotipo meno frequente, con conchiglia più grande e ornamentazione semplificata (macroconca) rispetto ai maschi. Questi ultimi avrebbero una conchiglia generalmente più piccola, (microconche), con ornamentazione più marcata, la cui imboccatura era sovente caratterizzata da strutture particolari sporgenti (rostro ventrale e apofisi laterali), con funzione ancora sconosciuta. Secondo questa ipotesi, i maschi sarebbero stati molto più numerosi delle femmine. Di fatto, non vi è alcuna evidenza nel materiale fossile su quali fossero i maschi e quali le femmine, e gli analoghi attuali (cefalopodi nautiloidi e coleoidi) hanno stili di vita e caratteristiche generali troppo diversi rispetto agli ammonoidi per costituire riferimenti accettabili integralmente.
La conchiglia delle ammoniti ha in generale la forma di una spirale avvolta su un piano (sebbene alcune specie, dette eteromorfe, abbiano un avvolgimento più complesso e tridimensionale) ed è proprio questa caratteristica ad aver determinato il loro nome. L'aspetto di questi animali, infatti, ricorda vagamente quello di un corno arrotolato, come quello di un montone (il dio egizio Amon era comunemente raffigurato come un uomo con corna di montone). Il celebre studioso romano Plinio il Vecchio (autore del trattato Naturalis Historia) definì i fossili di questi animali ammonis cornua, "corni di Ammone". Spesso il nome delle specie di Ammoniti termina in ceras, vocabolo greco (κέρας) il cui significato è, appunto, "corno" (p.es. Pleuroceras che etimologicamente significa corno con le coste). Le ammoniti sono considerate i fossili per eccellenza, tanto da essere spesso utilizzati come simbolo grafico della paleontologia.
-
Forma della conchiglia
La conchiglia delle ammoniti ha in generale la forma di una spirale avvolta su un piano (sebbene alcune specie, dette eteromorfe, abbiano un avvolgimento più complesso e tridimensionale) ed è proprio questa caratteristica ad aver determinato il loro nome.

L'immagine mostra le principali forme della conchiglia delle ammoniti. Sono comprese anche le forme più comuni tra quelle "eteromorfe", con l'avvertenza che in quest'ultimo caso l'elenco è ben lungi dall'essere esaustivo.
-
Area ventrale

La morfologia della regione ventrale è ugualmente importante dal punto di vista diagnostico: la forma generale e la presenza di carene e/o solchi (questi sono elementi che riguardano la morfologia della conchiglia e non fanno parte dell'ornamentazione) sono caratteri diagnostici spesso anche al livello di specie.
-
Peristoma
La camera d'abitazione è l'ultima camera, non settata, in cui vive l'organismo. Di lunghezza piuttosto variabile, è nella maggior parte delle forme compresa tra circa mezzo giro e un giro intero. Il bordo dell'apertura della camera d'abitazione si definisce peristoma. Molte ammoniti hanno peristomi semplici e lineari; tuttavia, nella maggior parte delle forme il peristoma è però più complesso: può essere delimitato da un rigonfiamento delle parete oppure preceduto da una contrazione; può essere dotato di orecchiette o apofisi laterali, mentre il ventre può essere dotato di un seno iponomico o al contrario di una proiezione anteriore, il rostro (vedi immagine). Non si conosce la funzione di queste strutture: secondo le due ipotesi prevalenti tra i ricercatori si tratterebbe di caratteri sessuali secondari o strutture difensive.
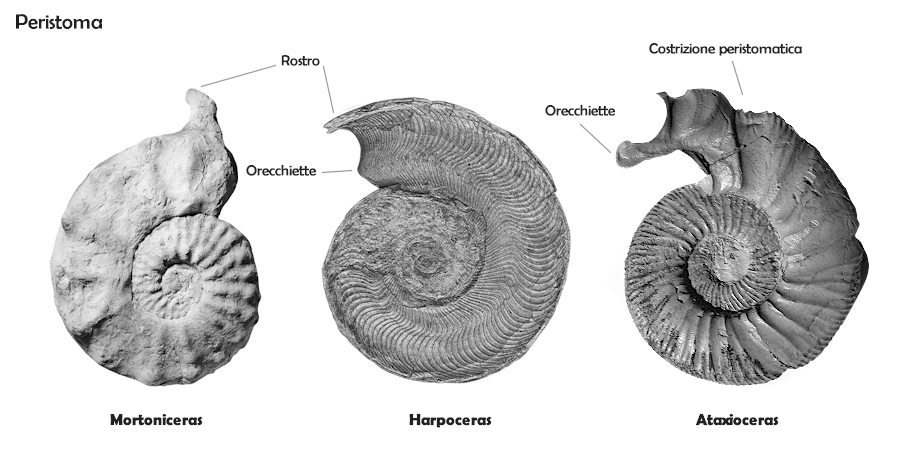
I peristomi successivi in diverse forme lasciano traccia sulla superficie della conchiglia sotto forma di costrizioni o varici: "restringimenti" locali del diametro della conchiglia che seguono l'andamento del peristoma (vedi immagine sopra). Queste costrizioni (che sono elementi morfologici e non fanno parte dell'ornamentazione) possono essere a seconda dei gruppi in numero che va da tre a una decina per giro, e rimarcano gli stadi di crescita dell'organismo. Talora sono solamente nei giri interni o in quelli esterni, e possono quindi variare con l'ontogenesi; in altri casi possono essere visibili solo nella parte ventrale o in quella periombelicale del giro.
Aptici
Come gli attuali nautili, molte ammoniti erano probabilmente in grado di ritirare completamente il proprio corpo all'interno della camera di abitazione della conchiglia; perciò avevano sviluppato strutture opercolari con le quali potevano chiudere l'apertura. Le forme più comuni sono costituite da due placche simmetriche, definiti aptici (aptychii). Sono stati rinvenuti anche opercoli singoli (non in coppia), che vengono definiti anaptici (anaptychus).
Gli aptici sono placchette di forma approssimativamente triangolare o sub-ovale, con un lato retto e il lato più corto curvilineo: assomigliano complessivamente a due valve di un lamellibranco (e in effetti, originariamente, prima che se ne evidenziasse la stretta associazione con le ammoniti, erano stati classificati come conchiglie di bivalvi o gusci di crostacei tipo ostracodi o fillocaridi). I due aptici hanno dimensioni identiche e sono speculari: come si vede dalla figura a fianco, uniti per il lato retto si adattano perfettamente all'apertura della conchiglia di un'ammonite. Talora le due "valve" sono saldate insieme, e in tal caso si ha un sinaptico (synaptychus). Gli aptici si trovano solamente in ammoniti del Mesozoico, solitamente isolati dalla conchiglia originale e talora in buono stato di conservazione.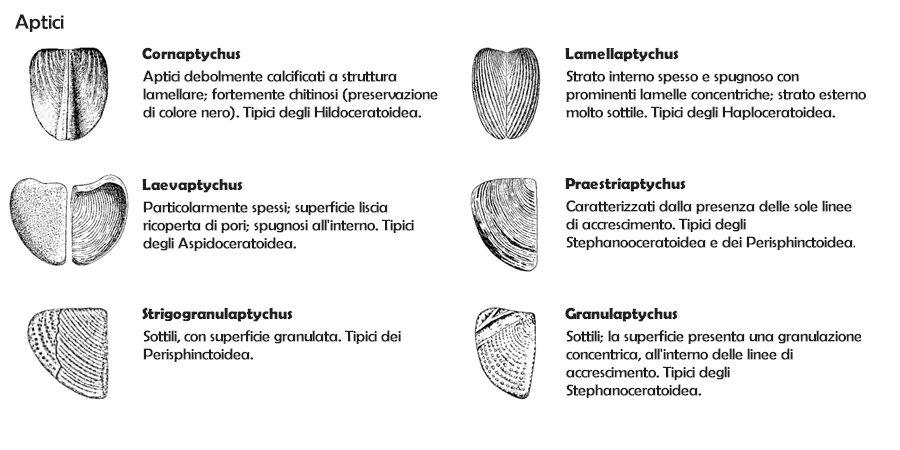
L'interpretazione della funzione degli aptici come opercoli è stata però messa in discussione a più riprese. Vari studi nell'ultimo trentennio suggerirebbero infatti la possibilità che gli aptici potessero essere parte di un particolare apparato mandibolare. Talora infatti, gli aptici sono stati rinvenuti associati ad un terzo elemento simile alla parte superiore della mascella di un nautiloide, e sono stati interpretati come elemento inferiore dell'apparato mandibolare. I problemi principali di questa interpretazione sono evidentemente le dimensioni piuttosto "ingombranti" degli aptici come mandibole, e il fatto che in posizione verticale occupano praticamente l'intera apertura della conchiglia.
Secondo le tendenze più recenti della ricerca in questo campo, però, le due interpretazioni non si escluderebbero a vicenda (gli aptici avrebbero avuto quindi la doppia funzione di opercoli e strutture nutritive). Gli aptici infatti si rinvengono sovente verso la parte ventrale della camera d'abitazione, in posizione coricata od obliqua: in tale posizione il margine interno (il lato più corto) degli aptici avrebbe potuto effettivamente esercitare una azione di "raschiamento" contro la parte superiore dell'apparato mandibolare, oppure di scavo. -
Ornamentazione

L'ornamentazione della conchiglia è costituita dagli elementi in rilievo, e può essere di tipo molto vario. In diversi casi (soprattutto nel Paleozoico), la conchiglia può essere liscia, e in tal caso mostra solo la presenza delle linee di accrescimento (come nei nautiloidi attuali). Generalmente nel Mesozoico (a partire dal Triassico) si sviluppano ornamentazioni sempre più complesse, soprattutto a decorso radiale:
- coste: elementi sporgenti a piega stretta e allungata, sviluppati radialmente;
- costellae: elementi radiali a tessitura molto fine;
- nodi: elementi sporgenti conici a tessitura fine (millimetrica);
- tubercoli: elementi sporgenti conici robusti (da millimetrici a centimetrici);
- bullae: elementi sporgenti, inflati, allungati perpendicolarmente alla direzione del giro, che spesso danno origine a fasci di coste o costellae;
- clavi: elementi sporgenti allungati parallelamente alla direzione del giro;
- spine: elementi sporgenti appuntiti e più o meno allungati, generalmente sottili; per elementi appuntiti robusti si usa talvolta il termine corni. Spesso le spine si originano da tubercoli, che ne costituiscono la base.
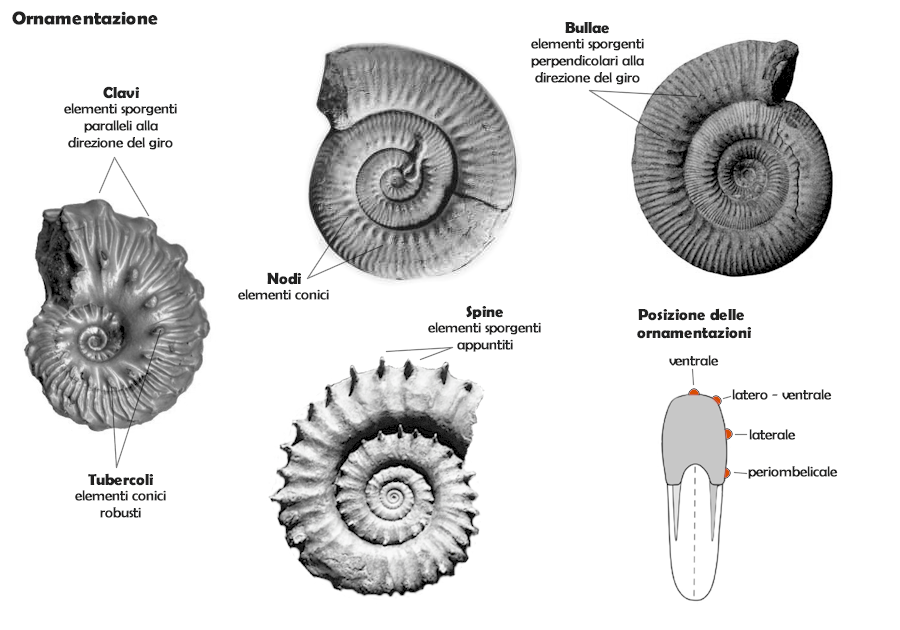
Tutti questi elementi possono essere variamente combinati, e frequentemente variano anche con la crescita dell'organismo, cioè con l'ontogenesi (in questo caso lo stadio adulto ha un'ornamentazione diversa rispetto agli stadi giovanili). La posizione degli elementi relativamente al giro della conchiglia può essere:
- laterale (sui fianchi);
- latero-ventrale (in prossimità o sul margine ventrale);
- periombelicale (in prossimità o sul margine ombelicale);
- ventrale (sul ventre); non rientrano in questa categoria solchi e carene, che fanno parte degli elementi morfologici della conchiglia.
L'ornamentazione è uno dei caratteri maggiormente diagnostici della conchiglia per le ammoniti, essendo sovente molto varia e sviluppata, e potendo variare rapidamente con l'evoluzione all'interno di uno stesso gruppo. -
Sutura
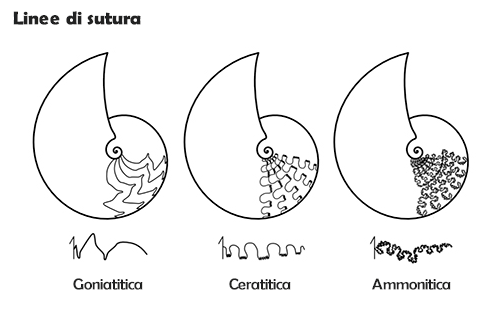
L'intersezione dei setti con la superficie interna della parete della conchiglia avviene lungo una linea, definita linea lobale, linea di sutura o semplicemente sutura, che nelle ammoniti è solitamente complessa (al contrario di quanto avviene nei nautiloidi, nei quali è di solito semplice: dritta o debolmente ondulata). L'andamento della linea di sutura era determinato da quello dei setti, che a sua volta era dovuto ad introflessioni ed estroflessioni del mantello presenti durante la fase di costruzione dei setti stessi, ed ha un decorso diverso a seconda dei gruppi tassonomici. È in effetti un carattere altamente diagnostico per la classificazione. Questo carattere è visibile ovviamente solo negli esemplari fossili privi della parete esterna, cioè sui modelli interni.
Se ne distinguono tre tipi fondamentali:
- sutura goniatitica: lobi e selle lineari, tipicamente non frastagliati, privi cioè di suddivisioni secondarie (eccezionalmente il lobo ventrale può presentare denticolazioni); selle da arrotondate ad angolose e lobi generalmente angolosi; caratterizzata nelle forme più primitive da pochi elementi, che tendono ad aumentare di numero nelle forme avanzate;
- sutura ceratitica: selle lineari e lobi con denticolazioni; da pochi elementi nelle forme più primitive a numerosi e relativamente complessi nelle forme evolute;
- sutura ammonitica: selle e lobi denticolati o frastagliati, spesso suddivisi in elementi accessori; un tipo particolare di sutura ammonitica è la sutura filloide, con selle suddivise in elementi più o meno numerosi a forma di foglia, con un elevato numero di lobi ombelicali (fino a sei) e una geometria di tipo frattale.
Infine, alcune famiglie di Ammonitida nel Cretaceo Superiore tendono a semplificare le suture fino ad una configurazione di tipo goniatitico o ceratitico (ad esempio la famiglia Tissotiidae e la sottofamiglia Mammitinae), con selle lineari emisferiche e lobi appuntiti e frastagliati. In questo caso si parla più propriamente di sutura pseudo-ceratitica, poiché non vi è alcun legame filetico diretto con le forme triassiche, dal momento che nel Giurassico sono conosciute solo forme con sutura ammonitica.
-
Indici morfometrici
La spirale descritta dall'avvolgimento della conchiglia può essere stretta (e in tal caso la conchiglia si dice involuta), oppure ampia (in tal caso si definirà evoluta). Un criterio largamente utilizzato dal punto di vista morfometrico è costituito dal rapporto tra altezza del giro e larghezza dell'ombelico (con rapporto >1 la conchiglia è involuta; se il rapporto è <1 la conchiglia è evoluta).
Generalmente, le ammoniti a conchiglia evoluta presentano uno scarso ricoprimento dei giri e quindi una regione ombelicale ampia, mentre quelle a conchiglia involuta per il motivo opposto presentano una regione ombelicale stretta e profonda (in alcuni casi l'ombelico può essere puntiforme o invisibile perché ricoperto da una callosità, come nell'attuale Nautilus pompilius).
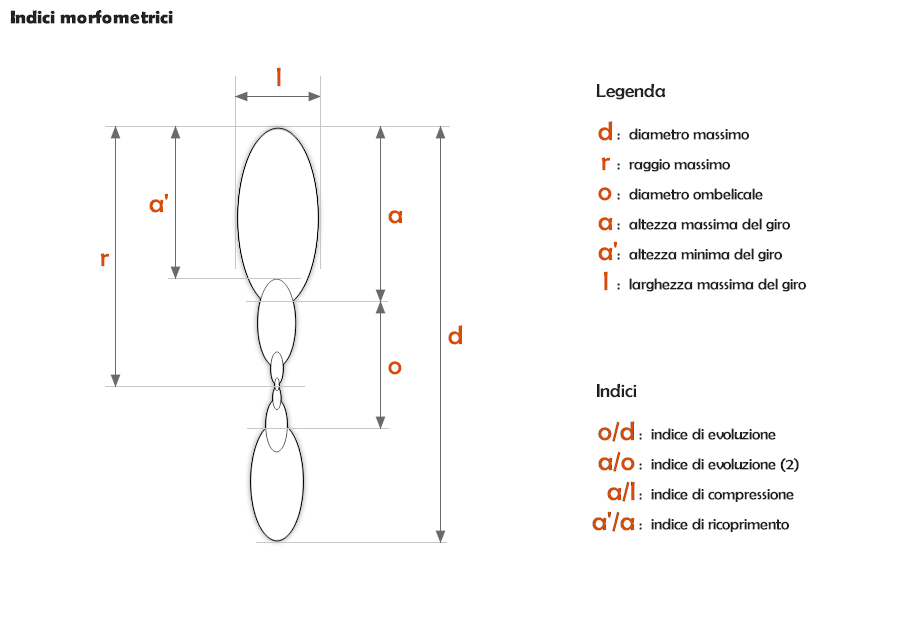
Un altro criterio morfometrico molto seguito per la definizione del grado di involuzione della conchiglia è quindi il grado di ricoprimento dei giri successivi: questo si può esprimere come percentuale di ricoprimento (data dal rapporto tra l'altezza totale del giro e la parte ricoperta, espresso in percentuale), oppure come rapporto tra la parte di altezza ricoperta e la parte libera. Questi parametri sono diagnostici per la classificazione.
Altri elementi morfometrici diagnostici sono la larghezza massima e l'altezza massima del giro (si utilizza il loro rapporto), che danno il grado di compressione del giro della conchiglia:
- giro compresso: altezza massima > larghezza massima
- giro depresso: larghezza massima > altezza massima
Occorre precisare che il grado di compressione del giro non determina necessariamente l'aspetto compresso o depresso della conchiglia nel suo insieme, che è influenzato anche dal grado di involuzione: esistono infatti conchiglie con giro depresso o sub-quadrato che hanno un aspetto decisamente compresso. Un esempio classico è il genere Perisphinctes, con giro sub-quadrato ma fortemente evoluto, e quindi con aspetto appiattito se vista sul lato ventrale (ma è scorretto definire questa conchiglia "compressa").
Quindi, la forma della conchiglia delle ammoniti viene complessivamente definita viene definita in base a diversi parametri:
- grado di involuzione
- grado di compressione
- forma della sezione del giro (che dipende dal tipo di curva generatrice)
- angolo di apertura del cono teorico
- piano (o piani) di avvolgimento
-
Fossilizzazione
Il processo di fossilizzazione inizia nel momento della morte dell'organismo. Una volta che giace sul terreno, o sul fondo di un lago o del mare, esso viene ricoperto di sedimento, e le parti molli cominciano a decomporsi. Gradualmente, i minerali presenti nelle acque percolanti sostituiscono quelli del guscio.
Solo una piccola percentuale di organismi riesce a fossilizzare, sia per le proprie caratterisiche (parti dure, come gusci, ossa o altro), sia per le condizioni di fossilizzazione (ambiente di deposizione, correnti, presenza di predatori, ecc.). In generale, l'ambiente subacqueo è molto migliore in quanto sede di deposizione, mentre l'ambiente subaereo è tipicamente sede di erosione.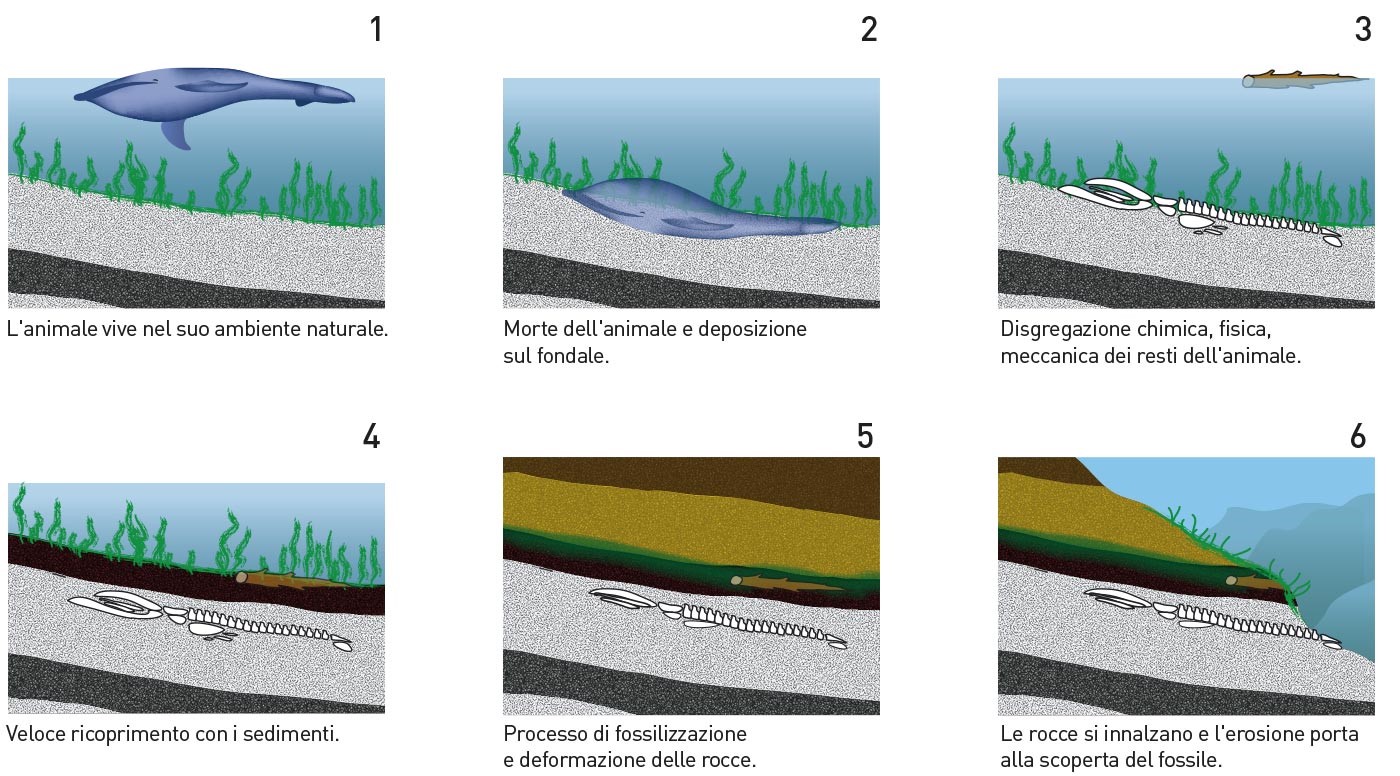
Le condizioni ideali di fossilizzazione sono:
- Il rapido seppellimento dei resti animali da parte del sedimento. Ciò previene l'azione dei predatori e dei batteri;
- Il rapido seppellimento nella cenere vulcanica. Molte ossa di dinosauro nell'ovest degli USA e in Cina sono stati rinvenuti sepolti nella cenere vulcanica;
- La presenza di parti dure nell'organismo (ad esempio, denti, ossa, gusci, legno);
- Condizioni di temperatura costante;
- Acque percolanti fortemente mineralizzate;
- Sedimentazione fine, migliore di quella più grossolana;
- Condizioni dell'ambiente a bassa energia, tali da impedire la rottura dei resti dell'organismo (ad esempio, per l'azione delle correnti o delle onde).
Tipi di fossilizzazione
Vi sono vari tipi di fossilizzazione, che dipendono strettamente dall'ambiente in cui si depositano i resti dell'organismo.
Il più comune è senz'altro il modello interno, che si forma quando il sedimento riempie le cavità dell'animale in precedenza sede delle parti molli. Spesso il guscio si dissolve, lasciando solo il calco del fossile, ma a volte il guscio si mineralizza e viene preservato.
Altre volte, durante il processo di fossilizzazione, le cavità vengono riempite dai minerali presenti nella acque percolanti (tipicamente carbonato di calcio), che dà luogo a bellissimi calchi spesso colorati. In questo caso si parla di ammonite permineralizzata
In ambiente riducente, con ossigenazione scarsa o assente, i gusci vengono rimpiazzati da pirite. Questa è molto instabile a condizioni normali, e le ammoniti piritizzate sono soggette a disgregarsi col tempo, se non trattate adeguatamente per resistere.
Si hanno poi fossili silicizzati o fosfatizzati, a seconda del contenuto minerale delle acque.
Vi sono altri tipi di fossilizzazione, che però non riguardano le ammoniti. Ad esempio, la conservazione in ambra; la resina di alcuni alberi e piante può invischiare insetti, ragni ed altri piccoli organismi. La resina poi indurisce e col tempo diviene ambra, preservando gli organismi in essa intrappolati.
Vi è poi l'incrostazione (ad esempio, le foglie nel travertino), la carbonificazione, ed altre estremamente rare e particolari (cerificazione, mummificazione, salificazione, distillazione).
-
Ecologia delle ammoniti
Come risulta da dati sia paleontologici che sedimentologici, la maggior parte delle ammoniti viveva in acque marine a salinità normale (30000-50000 ppm), in contesti di piattaforma continentale e di mare epicontinentale, in un intervallo batimetrico stimabile da zero a 200 metri di profondità. I fossili di ammoniti sono invece rari nei depositi di piattaforma carbonatica e nei sedimenti marini litorali e sono generalmente assenti in facies transizionali, come i delta e le piane di marea, ambienti caratterizzati da ampie e rapide variazioni di salinità. Sicuramente vi erano anche forme pelagiche adattate alle acque superficiali oceaniche.
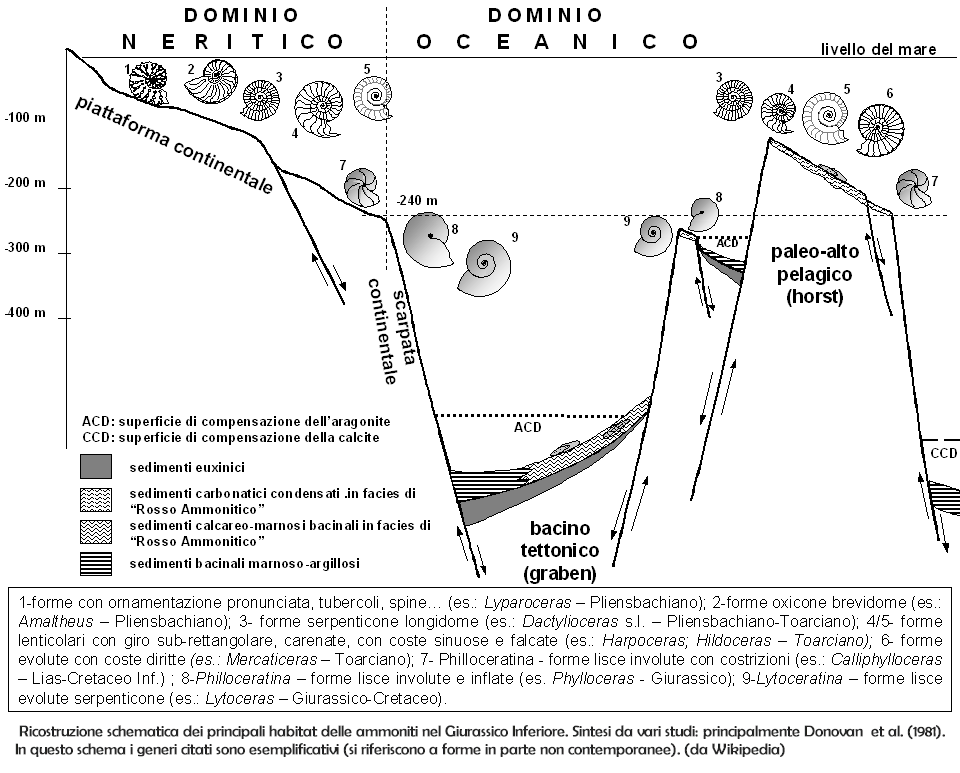
La grande varietà di morfologie comparse nella storia evolutiva di questo gruppo indica adattamenti a stili di vita molto diversi, che sono tuttora oggetto di dibattito tra gli specialisti. Da un punto di vista generale, morfologie appiattite e carenate, con profilo acuto, corrispondevano ad abitudini di vita nectoniche, da nuotatori relativamente veloci, mentre le forme più tondeggianti o globose, come anche le forme con ornamentazione molto sviluppata (meno idrodinamiche), sembrano essere state meno mobili, anche se queste considerazioni non dovrebbero essere eccessivamente generalizzate. Spesso, inoltre, durante l'ontogenesi i caratteri della conchiglia cambiano notevolmente, indicando una variazione di habitat significativa dagli esemplari giovanili agli adulti. Le forme "svolte" o con avvolgimento irregolare, infine, sono considerate dalla maggior parte degli studiosi come sicuramente non nectoniche, per lo meno nell'adulto: il dibattito verte soprattutto sulla loro caratterizzazione bentonica o planctonica.
-
Valore stratigrafico
Le ammoniti sono ottimi fossili fossili guida tutto il Mesozoico. Molte specie di ammoniti si sono evolute ed hanno velocemente compiuto il loro corso, durando da trecentomila anni ad alcuni milioni di anni. A causa della loro rapida evoluzione e dell'ampio areale, le ammoniti risultano essere un prezioso strumento per i geologi e paleontologi nei rilevamenti biostratigrafici per la datazione delle rocce sedimentarie, permettendo di collegare gli strati di roccia ad una precisa età geologica e di tracciare correlazioni tra sedimenti di aree diverse, anche a scala mondiale.
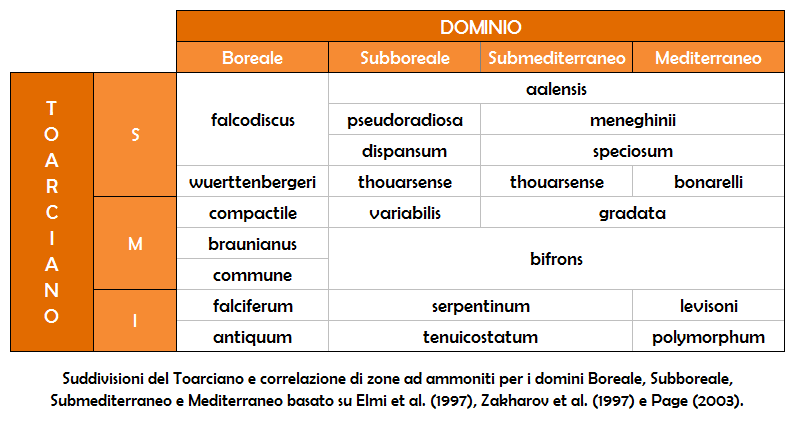
Le ammoniti offrono generalmente uno strumento stratigrafico di notevole precisione, pur tuttavia con alcuni limiti che non vanno sottovalutati. In particolare, le ammoniti sono spesso caratterizzate da evoluzione iterativa, cioè forme molto simili si possono ritrovare in tempi diversi e in gruppi non necessariamente relazionati, e su materiale fossile che non sia molto ben conservato spesso la determinazione della specie dell'indice di zona (e talora anche del genere) non è facile. L'indice di zona può essere anche assente o molto raro, per ragioni paleo-ecologiche o di distribuzione geografica.
Quindi sovente è l'associazione faunistica nel suo insieme a permettere la determinazione corretta della forma indice di biozona, o la stessa assegnazione alla biozona (e di conseguenza la datazione). Inoltre, per molte specie i caratteri tipici si vedono solo nella conchiglia adulta, pertanto sono necessari esemplari adulti e completi di camera di abitazione: per contro, molto spesso il paleontologo e lo stratigrafo hanno a che fare con materiale scarso e mal conservato. Ancora, (e questa è una regola che vale per tutti i gruppi faunistici), la documentazione fossile è raramente continua per un tempo geologico molto esteso, a causa della variazione laterale e verticale degli ambienti sedimentari: vi sono quindi lacune più o meno estese a scala locale che rendono spesso difficile la ricostruzione di un quadro stratigrafico completo e coerente.